| ESPERMIOHISTOGÉNESIS
Y ESPERMIACIÓN
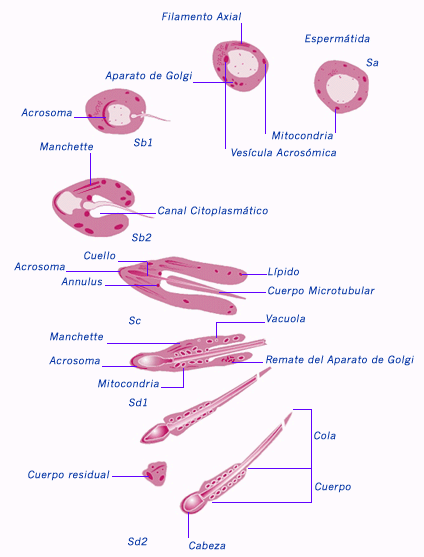
Las espermátidas son pequeñas y haploides (8 mm de diámetro), derivan de una espermatogonia pálida del tipo A y están unidas por puentes citoplasmáticos. Se disponen en pequeños conglomerados de ubicación cercana a la luz del túbulo seminífero. Estas células presentan RER abundante, mitocondria abundante y complejo de Golgi bien desarrollado. Durante la transformación, acumulan enzimas hidrolíticas, ordenan los organelos y los reducen en número, forman el flagelo y un citoesqueleto asociado, con reducción de citoplasma. Este proceso de espermiohistogénesis se subdividió en cuatro fases: fase de Golgi, fase de capucha o cubierta, fase acrosómica y fase de maduración. La característica principal de este proceso se relaciona con la formación del acrosoma desde el complejo de Golgi, condensación y elongación del núcleo, formación de un flagelo movible, y un desprendimiento extensivo de citoplasma. Los factores responsables de este cambio nuclear y citoplasmático son todavía poco conocidos. Fase de Golgi: las enzimas hidrolíticas formadas en el RER, se modifican en el aparato de Golgi y empacan en la cara trans de él, como pequeños gránulos proacrosómicos. La fase de Golgi comienza cuando las espermátidas avanzan hacia el espermatocito II. Se observan gránulos proacrosómicos que coalescen dentro de la vesícula acrosómica recubierta por membrana, formando un sólo gránulo largo, el gránulo acrosómico, que rápidamente se dirige a la región perinuclear. La posición de la región acrosómica en el núcleo, identifica el polo anterior de la espermátida. Paralelamente, los centríolos se alejan del núcleo y uno de ellos participa en la formación del axonema flagelar. Iniciada la generación de microtúbulos, los centríolos regresan a la posición perinuclear y participan en la formación de la pieza intermedia. Fase de capucha o cubierta: Durante esta fase, la vesícula acrosómica aumenta de tamaño y su membrana rodea parcialmente al núcleo. Cuando esta vesícula alcanza su tamaño final, se identifica como acrosoma (cubierta acrosómica). Los centríolos se visualizan como proximal, en el polo posterior del núcleo, y dista, en el curpúsculo del flagelo. Durante esta fase, el acrosoma que se desarrolla desde la vesícula acrosómica se extiende para cubrir parcialmente la cara anterior del núcleo. El acrosoma presenta una membrana acrosómica interna y otra externa encerrando el contenido acrosómico. Entre el ambiente interno y la membrana acrosómica interna, se forma un material filamentoso granular. Además, en la región del acrosoma, la carioteca nuclear pierde los poros nucleares y aparece denso, posiblemente por la condensación de la cromatina. Fase acrosómica: Esta fase se caracteriza por la presencia de varias modificaciones en la morfología de la célula. El núcleo se condensa, la célula se alarga y las mitocondrias cambian su ubicación. Los cromosomas se condensan y, conforme disminuye su tamaño, el núcleo se aplana y adquiere la forma característica. La característica principal de esta fase es la orientación del polo anterior del núcleo de la espermátida dirigido hacia la base de los túbulos seminíferos, y la elongación y condensación nuclear. El citoplasma se desplaza hacia la región luminal del túbulo seminífero. De esta manera, la región acrosómica se acerca rápidamente a la membrana plasmática y la célula se alarga. El núcleo aumenta de volumen y se alarga, mientras los gránulos de cromatina también se alargan y se vuelven uniformes en tamaño y dispersión, finalmente el núcleo adquiere una apariencia homogénea, con una coloración oscura, desprovisto de estructuras internas. Cuando el acrosoma cesa de crecer el complejo de Golgi se separa del polo anterior del núcleo y migra libremente en él. Al mismo tiempo, el ensamblaje de microtúbulos citoplasmáticos permite la formación de una vaina cilíndrica que se une al polo caudal del núcleo, cerca del margen posterior de la capa acrosómica. Esta vaina es llamada el machete, o tubo caudal y su función es desconocida. Cerca de los centríolos se forma una estructura con forma de anillo, que migra bajo el flagelo. Las mitocondrias se alinean en la región del cuello de la estructura. Esta porción del cuello es llamada la pieza media. Cuando la migración mitocondrial cesa, el tubo caudal o machete desaparece. Distal a la pieza media se desarrolla una vaina fibrosa, las nueve fibras longitudinales. Fase de maduración: se caracteriza por la pérdida de citoplasma por parte de la espermátida. Conforme se libera el exceso de citoplasma, se forman los espermatozoides individuales, que serán liberados desde las células de Sertoli. Las células de Sertoli fagocitan los residuos citoplasmáticos, y los espermatozoides libres pasan hacia la luz del túbulo seminífero (espermiación). Los espermatozoides recién formados no tienen movilidad y no pueden fecundar al oocito. Sólo después de la maduración en el conducto epidimario adquieren movilidad progresiva. ESPERMIACIÓN Proceso fisiológico de liberación de los espermatozoides a la luz tubular, regulado por la célula de Sertoli y bajo la influencia de la hormona LH, cuyo efecto sería el de facilitar el transporte de agua y sodio al espacio intracelular, fenómeno del cual dependen los cambios estructurales conducentes a la liberación. En algunos mamíferos se produce en forma espontánea, mientras que en otros es por el estímulo coital. |